
First record of the ferret-badger Melogale cucphuongensis Nadler et al.,2011(Carnivora:Mustelidae),with description of a new subspecies,in southeastern China
2019-10-31 10:51:10SongLi,Guo-HuaYu,ShuoLiu等
Zoological Research 2019年6期
DEAR EDITOR,The ferret-badger Melogale cucphuongensis was first described from Cuc Phuong National Park in Vietnam,with no subsequent reports in later years.During our surveys of the Wuyishan Mountains of Fujian Province in southeastern China during May 2018, a Melogale specimen was identified.Analysis based on pelage and skull characteristics as well as molecular data indicated it to be a new subspecies,which we nominated as Melogale cucphuongensis guadunensis subsp.nov.This is the first record of the species in China.Therefore,this research not only expands the distribution range of the species beyond Vietnam,but also indicates that geographic variation of the species should have been proceeding in southeastern China.
Ferret-badgers(Melogale I.Geoffroy,1831)were originally assigned into four species distributed in Indochina,Java,Bali,China,and northeastern Borneo(Corbet&Hill,1992;Wilson&Reeder,2005).Among them,Melogale everetti Thomas,1895 is restricted to Mount Kina Balu in northern Borneo,M.orientalis Horsfield,1821 occurs in Indonesia,M.personata Geoffroy,1831 is distributed from the southern Indo-China Peninsula northward to Assam,including southwestern China,and M. moschata Gray, 1831 is found in China (from southwestern to eastern areas,including Hainan and Taiwan),India,Laos,and Vietnam(Corbet&Hill,1992;Smith&Xie,2009;Wang,2003;Wilson&Reeder,2005).Currently,there are only two Melogale species, M. personata and M.moschata,distributed in China.Based on skull morphology,pelage characteristics,and mitochondrial cyt b gene analyses,Nadler et al. (2011) described a new species (M.cucphuongensis) from Vietnam, which occurs sympatrically with M.moschata and M.personata.However,no subsequent reports on these or new species have occurred in recent years.
We conducted a mammal survey in the Wuyishan Mountains of southeastern China in May 2018.We discovered one male Melogale individual(collection number:2018000079)with a crushed head,which had died on a road near Guadun village(E177.64°,N27.73°,Altitude:1074 m a.s.l.).
We described the pelage and dentition characteristics(despite the damaged skull)and carried out morphological measurements. In total, four cranial measurements were taken with a digital caliper to an accuracy of 0.01 mm,including:upper tooth row(UTR),lower tooth row(LTR),P4:length of P4,P1-M1:length from front edge of P1 to end of M1.Body weight(BW)and four external measurements were also taken,including:body length(BL),tail length(TL),hind foot length(HF),and ear length(EL).
Total genomic DNA was extracted from muscle tissue collected from the specimen.The tissue sample was digested using proteinase K, and subsequently purified following standard phenol/chloroform isolation and ethanol precipitation.A fragment encoding the complete cyt b gene was amplified using primer pairs L14724:5'-CGAAGCTTGATATGAAAAACC ATCGTTG-3',and H15915:5'-AACTGCAGTCATCTCCGGTT TACAAGAC-3'(Irwin et al.,1991).Polymerase chain reaction(PCR)amplification was performed in 50 μL reactions under the following cycling conditions:initial denaturing step at 95°C for 4 min;35 cycles of denaturing at 94°C for 60 s,annealing at 51°C for 60 s,extension at 72°C for 60 s;and final extension at 72°C for 10 min.Sequencing was conducted using the corresponding PCR primers,with the new sequence deposited in GenBank under Accession No. MK167045.Available homologous sequences of other species were obtained from GenBank.Taxidea taxus was selected as the outgroup.
Sequences were aligned using MUSCLE with default parameters in MEGA 7 (Kumar et al., 2016). Uncorrected pairwise distances were calculated in MEGA 7. The best substitution model was selected using the Akaike Information Criterion (AIC) in MODELTEST v3.7 (Posada & Crandall,1998). Bayesian inferences were performed in MRBAYES 3.1.2(Huelsenbeck&Ronquist,2001)based on the selected substitution models (GTR+I+G). Two runs were performed simultaneously with four Markov chains starting from a random tree.The chains were run for 3 000 000 generations and sampled every 100 generations.The first 25%of sampled trees were discarded as burn-in when the standard deviation of split frequencies of the two runs was less than 0.01.The remaining trees were then used to create a consensus tree and estimate Bayesian posterior probabilities(BPPs).
Taxonomy
Melogale cucphuongensis guadunensis Li et al.,subsp.nov.
Holotype:KIZ 2018000079,♂,adult,collected on 13 May 2018,from Guadun(E177.64°,N27.73°,Altitude:1074 m a.s.l.),Wuyishan,Fujian,China.The holotype is preserved at the Kunming Natural History Museum of Zoology, Kunming Institute of Zoology,Chinese Academy of Sciences,Yunnan,China.
Etymology:The new subspecies is named according to the type locality.
Diagnosis:Cheeks and rear of eye yellow-brown;edge of earconch yellow-brown;small whitish patch between eyes and on ears respective,and on the neck back also;tail pale brown all around with whitish hairs tips,transforming to yellow-brown at end.P4~1/3 length of P1-M1(length from front edge of P1to end of M1).
Description:Head brown-black,deeper in color than back;naked nose terminal elongated and wedge-like, mouth positioned clearly behind nose. Vibrissae dark brown and long, anteorbital area black-brown. Small whitish patch between eyes and at vertex between ears.Frontal part brown.Cheeks and rear of eye yellow-brown; edge of ear-conch yellow-brown(Figure 1).
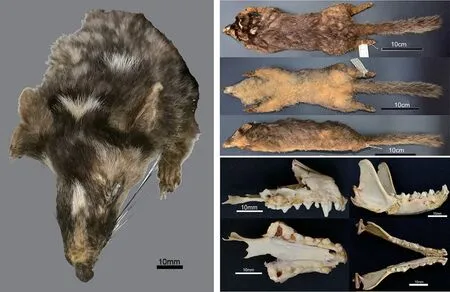
Figure 1 Holotype of Melogale cucphuongensis guadunensis Li et al.,subsp.nov.
General color of back brown with long frosted-like,whitish hair tips;whitish patch on center of back of neck,with no whitish stripe from center of occiput backward to neck and shoulders.Under part of body yellow-cream.Chin and throat thinner than abdomen(Figure 1).
Feet covered in brown hairs overall,but naked on soles;forefeet claws stronger than those of hindfeet and slightly curved.Tail bushy,pale brown all around with whitish hair tips,transforming to yellow-brown at end(Figure 1).
As noted above,although the skull was damaged,most teeth were preserved.Dental formula:I3/3,C1/1,P4/4,M1/2=38.C1developed,height twice that of P1;P1smallest cheek teeth;P2nearly same as P3,but less than latter more or less;P4well developed,mesostyle markedly larger than paracone,metacone elongated backward significantly, forming sharp dental ridge,outer edge slightly concaved,parastyle lingually more developed than protocone;M1developed,lingual edge extended to form oval,clear concavity between paracone and metacone at labial edge;Position of I2obviously backward;C1developed,with curving sharp rear rim;P1much smaller than P2and P3;P4less complicated than P4with developed center cusp;M1with developed anterior trigon and three similar size cusps on crown,second half without clear cusps,but with sharp edge;M2contract conspicuous,round,with outer and inner cusps(Figure 1).
Synonyms:Melogale moschata ferreogrisea (Hilzheimer,1905).
Measurements:Holotype measurements(mm)of Melogale cucphuongensis guadunensis Li et al.,subsp.nov.are listed in Table 1.
Molecular genetics:Based on a 1 140 bp fragment of cyt b and using Taxidea taxus as the outgroup(all obtained from GenBank except for holotype in the study), Bayesian phylogram of M. personata, M. moschata, M.cucphuongensis(JN032659 which is only 423 bp of cyt b)and Arctonyx eight genera of Mustelidae those who distribute in China are displayed. Each group is listed in the tree followed by its GenBank Accession No.As shown in Figure 2,M.c.guadunensis subsp.nov.and M.c.cucphuongensis formed a sister group,which,combined with the sister group of M. personata and M. moschata, formed the Melogale clade.
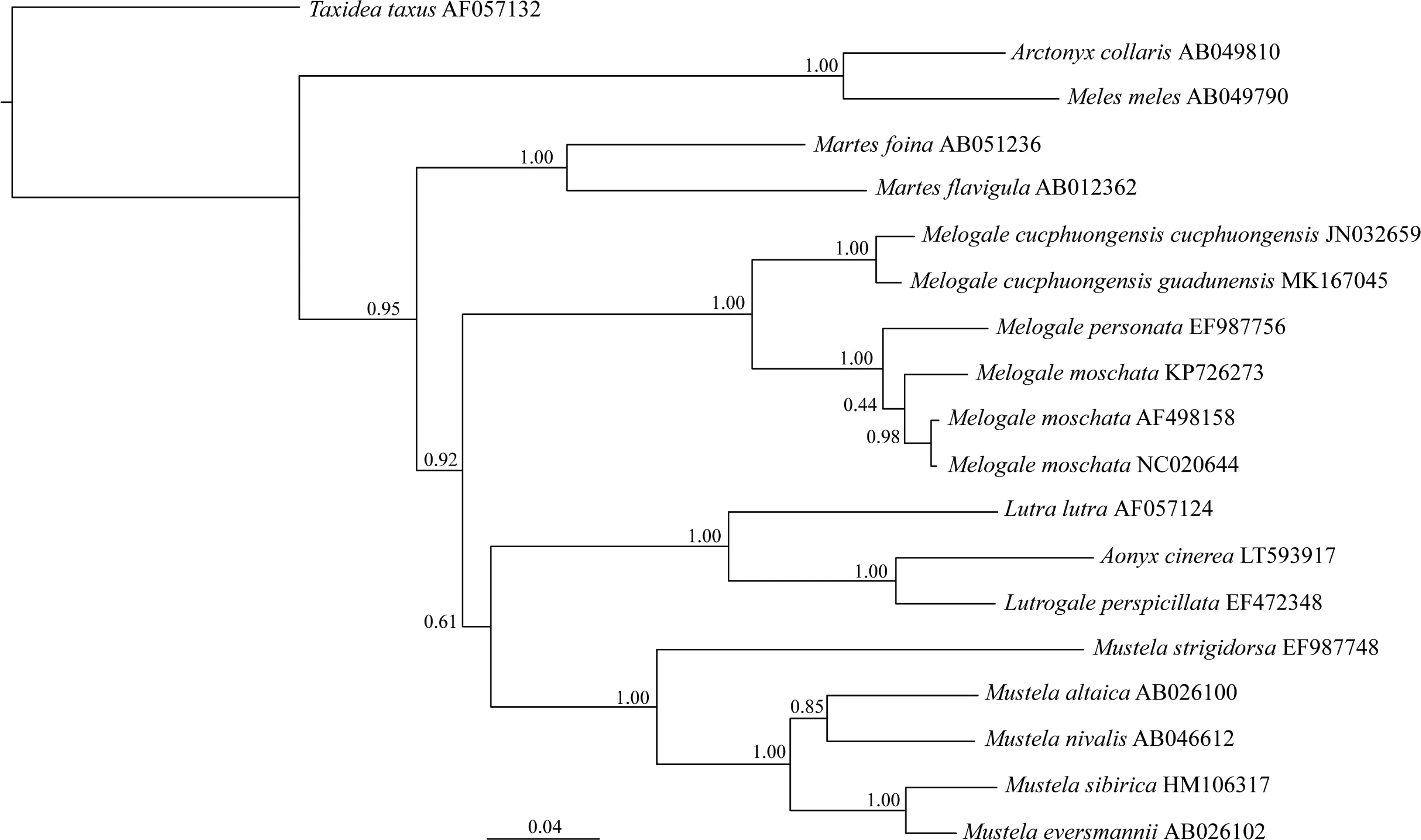
Figure 2 Bayesian phylogram inferred from 1 140 bp fragment of cyt b
Based on the uncorrected pairwise distances,the genetic difference between M.c.guadunensis subsp.nov.and M.moschata was 7.46-7.63, between M. moschata and M.personata was 4.12-4.82,between M.c.guadunensis subsp.nov. and M. personata was 8.42, between M. c.cucphuongensis and M.personata was 8.27,and between M.c.guadunensis subsp.nov.and M.c.cucphuongensis was 1.42(Table 2).
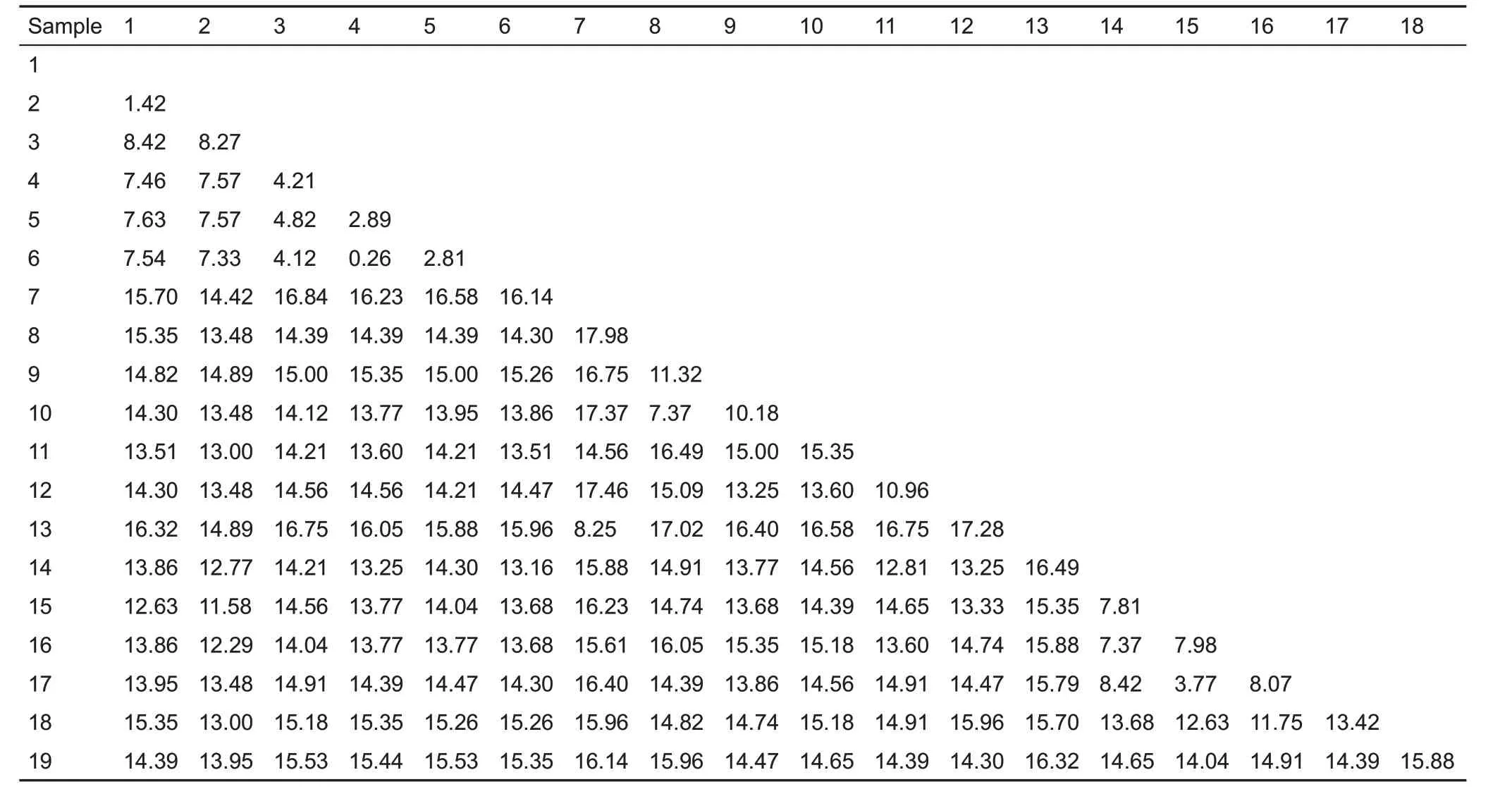
Table 2 Uncorrected pairwise distances estimated from a 1 140 bp fragment of cyt b
In contrast to the description of the characteristics of M.c.cucphuongensis(Nadler et al.,2011),we list a key to the subspecies below:
Key to Melogale cucphuongensis subspecies
1) Cheeks yellow-brown; tail pale brown all around with whitish hairs tips,transforming to yellow-brown at end;P4~1/3 length of P1-M1…M.c.guadunensis subsp.nov.
2) Cheeks brown,with no white markings;tail uniform in color;P4~1/4 length of P1-M1………M.c.cucphuongensis
Melogale c. guadunensis subsp. nov. is similar to M.moschata in a number of characteristics, including brownblack head(deeper in color than back),naked wedge-like nose terminal,and small whitish patch between eyes and ears, respectively (Figure 1). As M. moschata occurs throughout southern China(Corbet&Hill,1992;Smith&Xie,2009;Wang,2003;Wilson&Reeder,2005),many previously discovered Melogale samples from China, especially from central and southeastern areas,have long been regarded as M.moschata.However,M.c.guadunensis subsp.nov.can be clearly distinguished from M.moschata by the following characteristics: whitish stripe absent on center of occiput backward to neck(vs.present)(Figure 1);rhinarium wedgelike(vs.subcircular)(Figure 1);P4~1/3 length of P1-M1(vs.~1/4)(Table 1);tail length over 50%of body length(Table 1)(vs.less than 50%)(Gao,1987;Zheng&Xu,1983).
The genetic differences between M.c.guadunensis subsp.nov.and M.moschata and M.c.guadunensis subsp.nov.and M.personata are greater than that between M.moschata and M. personata. Furthermore, the genetic difference between M. c. guadunensis subsp. nov. and M. c.cucphuongensis is only 1.42(Table 2).These results not only confirm that M. c. guadunensis subsp. nov. is markedly different from M. moschata and M. personata, but also demonstrate that M.c.guadunensis subsp.nov.and M.c.cucphuongensis are the same species.Phylogenetic analysis also indicated that M.c.guadunensis subsp.nov.and M.c.cucphuongensis formed the M.cucphuongensis clade,and M.moschata and M.personata were phylogenetically closer than either to M.cucphuongensis(Figure 2).
Melogale c. cucphuongensis, which is known from Cuc Phuong National Park in Vietnam, is found in limestone formations with primary, slightly degraded primary, and secondary forests (Nadler et al., 2011). Melogale c.guadunensis subsp.nov.is distributed in Guadun,Wuyishan,Fujian,China,within subtropical evergreen broadleaf forest and coniferous and broad-leaved mixed forest.The two type localities are separated by the vast expanse of southern China, which raises the question: how does the species disperse across such an expansive area? Currently, very limited information is available in regard to its biology and ecology.Thus,additional studies on its distribution,population status,and biological habits are required.
NOMENCLATURAL ACTS REGISTRATION
The electronic version of this article in portable document format will represent a published work according to the International Commission on Zoological Nomenclature(ICZN),and hence the new names contained in the electronic version are effectively published under that Code from the electronic edition alone(see Articles 8.5-8.6 of the Code).This published work and the nomenclatural acts it contains have been registered in ZooBank,the online registration system for the ICZN.The ZooBank LSIDs(Life Science Identifiers)can be resolved and the associated information can be viewed through any standard web browser by appending the LSID to the prefixhttp://zoobank.org/.
Publication LSID:urn:lsid:zoobank.org:pub:6C7D10B8-8669-41D2-ACA5-DF16EAC58C27.
Nomenclatural act LSID:urn:lsid:zoobank.org:act:CFF1216E-445F-413A-B5DB-0AAA256F4135.
SCIENTIFIC FIELD SURVEY PERMISSION INFORMATION
Permission for field surveys in Wuyishan,Fujian,China was granted by Research Monitoring Center,Wuyishan National Park,and Fujian Wuyishan Forest Ecosystem Research Station.
COMPETING INTERESTS
The authors declare that they have no competing interests.
AUTHORS'CONTRIBUTIONS
S.L.designed the study and wrote the manuscript.G.H.Y.finished the genetic analyses.S.L.and C.S.J.performed the field surveys.All authors read and approved the final version of the manuscript.
ACKNOWLEDGEMENTS
This work was conducted at the Kunming Institute of Zoology,Chinese Academy of Sciences(CAS).We are grateful to the anonymous reviewers for their valuable comments and remarks regarding the manuscript.Special thanks are given to Mr.Xiao-Feng Ma for photography,and Mr.Kang Luo and Mr.Ming-Jin Pu for their hard work on specimen collection.

- Zoological Research的其它文章
- Diversity scaling of human vaginal microbial communities
- Direct sunlight exposure reduces hair cortisol levels in rhesus monkeys(Macaca mulatta)
- Ongoing green peafowl protection in China
- A new species of the genus Xenophrys(Anura:Megophryidae)from northern Thailand
- A new species of the genus Raorchestes(Anura:Rhacophoridae)from Yunnan Province,China
- Sinocyclocheilus sanxiaensis,a new blind fish from the Three Gorges of Yangtze River provides insights into speciation of Chinese cavefish