
A new species of Luganoia (Luganoiidae, Neopterygii)from the Middle Triassic Xingyi Biota, Guizhou, China
2020-11-11 12:59:42XUGuangHui
古脊椎動(dòng)物學(xué)報(bào)(中英文) 2020年4期
XU Guang-Hui
(1 Key Laboratory of Vertebrate Evolution and Human Origins of Chinese Academy of Sciences, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences Beijing 100044 xuguanghui@ivpp.ac.cn)
(2 CAS Center for Excellence in Life and Paleoenvironment Beijing 100044)
Abstract Neopterygii is the largest known group of ray-finned fishes today, and the Luganoiidae,recognized by its specialized skull and greatly deepened flank scales, is a stem lineage of this group in the Middle Triassic. Since its naming in 1939, the Luganoiidae has been represented solely by the marine species Luganoia lepidosteoides found near the Ladinian/Anisian boundary of the southern and eastern Switzerland, northern Italy and southeastern Spain. Here, I report the discovery of a new species of this genus, Luganoia fortuna sp. nov. based on a nearly complete specimen from the late Middle Triassic (Ladinian) marine deposits exposed near Xingyi,Guizhou, China. The discovery represents the first record of the Luganoiidae in Asia, indicating that the biogeographical distribution of this family is much wider than previously recognized.Comparative studies of the new species with the type species from Europe reveal some anatomical features previously unnoticed in Luganoia, e.g., the presence of antorbitals, absence of a platelike quadratojugal, presence of a narrow naked region of the body at the base of the dorsal fin, presence of a small anal fin closer to the caudal fin than to the pelvic fin, and absence of postcleithra. An amended diagnosis of the genus is presented. Moreover, the distinguishable features between the Chinese and European species are highlighted in this study, and they provide valuable information on the morphological diversification of Luganoia.
Key words Xingyi, Guizhou; Triassic; Luganoiidae; Neopterygii; osteology; paleobiogeography
LuganoiaBrough, 1939 is the type genus of the Luganoiidae, renowned by its specialized skull and greatly deepened flank scales. As a derived stem-neopterygian, the genus is of particular interest to paleoichthyologists for investigating the rise of the neopterygian crown(Patterson, 1973; Gardiner and Schaeffer, 1989; Xu et al., 2015a, 2018b; Xu and Ma, 2016;Wen et al., 2019). When it was named,Luganoiaoriginally included three species:Luganoia lepidosteoidesfrom the Middle Triassic of Besano, Lombardy in Italy and Monte San Giorgio,Tessin in Switzerland, andL.intermediaandL. macrodentatafrom the Middle Triassic of Besano, Lombardy in Italy (Brough, 1939). However, Bürgin (1992) suggested that the latter two are younger synonyms of the former based on his comparative studies of the type material originally described by Brough (1939) and new specimens from the Monte San Giorgio area. Later,L. lepidosteoideswas also found from the Middle Triassic of southeastern Spain(Cartanyà, 1999) and eastern Switzerland (Bürgin, 1999). In the same publication, Brough (1939)named the other luganoiid genusBensaniafrom the Middle Triassic of Besano, Lombardy in Italy, but Herzog and Bürgin’s (2005) studies showed that itis actually not a luganoiid but rather a crown-neopterygian. As thus,Luganoiais the only valid genus of the Luganoiidae.Traditional studies (Brough, 1939; Patterson, 1973; Bürgin, 1992) placed the Luganoiidae in the paraphyletic ‘Subholostei’ or ‘Perleidiformes’, but recent studies (Cartanyà, 1999; Xu et al.,2015a, 2018b; Wen et al., 2019) prefer to place the family in its own order, Luganoiiformes Lehman, 1958.
Here, I report the discovery of a new species ofLuganoiabased on a nearly complete specimen from the lower part of the Zhuganpo Member of the Falang Formation exposed in Wusha, Xingyi, Guizhou Province. The discovery represents the first record ofLuganoiaoutside of Europe, adding new information on the morphological diversification and biogeographical distribution of the genus. The fossil fish is well-preserved in the dark grey thin-bedded argillaceous limestone, permitting a detailed description of its morphology. Other macrofossils from the same fossiliferous horizons include invertebrates, other groups of rayfinned fishes (Su, 1959; Jin, 2001; Liu et al., 2002, 2003; Xu et al., 2012, 2015b, 2018a, b;Xu and Zhao, 2015; Tintori et al., 2015; Xu and Ma, 2018), a coelacanth (Geng et al, 2009),and diverse marine reptiles (Young, 1958; Li, 2006; Li and Jin, 2009; Li C et al., 2016; Lu et al., 2018). The whole fossil assemblage documents the Xingyi Fauna or Biota in the Triassic Yangtze Sea (a part of the eastern Paleotethys Ocean) in southern China (Benton et al., 2013;Lu et al., 2018). Biostratigraphical studies of invertebrates and marine reptiles (Young, 1958;Chen, 1985; Li and Jin, 2009; Zou et al., 2015) generally suggest a late Middle Triassic(Ladinian) age for this biota. Although a younger Late Triassic (Carnian) age was once proposed (Wang et al., 1998), this was based on inaccurate conodont identifications (Zou et al.,2015). A recent zircon U-Pb dating ((240.8 ± 1.8) Ma; Li Z G et al., 2016) further supports a late Middle Triassic (Ladinian) age for the Xingyi Biota.
1 Material and methods
The specimen is curated at the fossil collections of the Institute of Vertebrate Paleontology and Paleoanthropology (IVPP), Chinese Academy of Sciences in Beijing, China.It was mechanically prepared with sharp steel needles under an Olympus SZX7 microscope.For better contrast, the specimen was dusted with ammonium chloride (NH4Cl) before being photographed. The relative position of fins and scale counts were expressed following Westoll
(1944), in which the letters D, P, A, and C are followed by a particular numerical value to denote the number of vertical scale rows between the first complete row behind the pectoral girdle and the insertion of the dorsal (D), pelvic (P), anal (A), and caudal (C) fins, respectively.The letter T denotes the total number of scale rows between the pectoral girdle and the caudal inversion. The traditional actinopterygian nomenclature of Gardiner and Schaeffer (1989) and Grande and Bemis (1998) are generally followed, for ease of comparison with most existing literature.
2 Systematic paleontology
Neopterygii Regan, 1923 Luganoiiformes Lehman, 1958 Luganoiidae Brough, 1939 Luganoia Brough, 1939
Emended diagnosisLuganoid fishes distinguished from other stem-neoptergyians by the following features (possible autapomorphies identified with an asterisk): fusion of median rostral with premaxillae, bearing long and slender posterior process (*); frontals completely fused into single ossification and greatly constricted above orbital center (*); presence of paired parieto-dermopterotics; presence of three or four extrascapulars; fusion of lacrimal with maxilla; lacrimo-maxilla with large triangular dorsal process and concave oral margin,ending below orbital center (*); presence of multiple supraorbitals; separation of suborbitals into two sets by broad preopercle (*); preopercle with long, anteriorly tapering process contacting lacrimo-maxilla; presence of peg-like teeth only on anterior half of jaws; absence of postcleithrum; body naked near base of dorsal fin; caudal fin with truncate profile; presence of about ten epaxial rays in caudal fin; anal fin small, located closer to caudal fin than to pelvic fins; and single row of 13 greatly deepened lateral line scales contacting two or three horizontal rows of short scales posteriorly (*).
Type speciesLuganoia lepidosteoidesBrough, 1939.
DistributionLatestAnisian to Ladinian, Middle Triassic; Besano, Lombardy, Italy;Monte San Giorgio, Tessin and Davos, Graubünden, Switzerland; Alcover, Catalonia, Spain;Xingyi, Guizhou, China.
Luganoia fortunasp. nov.
(Figs. 1-4, 5A)
EtymologyThe specific epithet means lucky.
HolotypeIVPP V 25693, a nearly complete specimen with a dorsally crushed skull and a laterally compressed postcranium.
Locality and horizonXiemi, Wusha, Xingyi, Guizhou Province, China; lower part of Zhuganpo Member of Falang Formation, Ladinian (~240 Ma), Middle Triassic (Xu and Zhao,2015; Li Z G et al., 2016).DiagnosisA new species ofLuganoiadistinguished from the type species (L.lepidosteoides) by the following features: frontals 3.3 times as long as parieto-dermopterotics;three extrascapulars; subopercle 1.5 times as large as opercle, ornamented by many ridges on its ventral portion; single suborbital above preopercle; six or seven suborbitals anteroventral to preopercle and posterior to lacrimo-maxilla; preopercle with anteroventrally tapering ramus;presence of three peg-like teeth on rostro-premaxillae; three scales between posttemporals; 15 pectoral fin rays; 9 pelvic fin rays; 11 dorsal fin rays; 12 principal caudal fin rays; and pterygial formula of D20/P14, A24, C29/T33.
3 Description
General morphology and sizeThe general morphology ofLuganoia fortunasp. nov. is very similar to that of the type species (L. lepidosteoides), having a pointed snout, a fusiform body with greatly deepened lateral line scales in the anterior flank region, and an abbreviated heterocercal caudal fin (Fig. 1). The relatively large dorsal fin inserts slightly posterior to the origins of the pelvic fins. The small anal fin is located closer to the caudal fin than to the pelvic fins. The holotype (Fig. 1) has a head length of 21 mm, a standard length of 64 mm, and a total length of about 73 mm. The greatest body depth is 17 mm, located midway between the posterior margin of the opercle and the origin of the dorsal fin. Although small in size, this specimen is considered to be an adult individual because the skull-roofing bones are firmly sutured. In comparison, the new species is slightly larger thanL. lepidosteoides, which has a standard length of 50 mm (Bürgin, 1992).
SnoutAs a characterized feature of this genus, the rostral is fused with the premaxillae.The rostro-premaxillae has a nearly rectangular anterior plate and a long and slender posterior process that tapers posteriorly and contacts the fused frontals with a pointed end (Fig. 2). The rostro-premaxillae is 0.74 times as long as the fused frontals. Three teeth are present at the oral margin of the anterior plate of the rostro-premaxillae; they are strong, peg-like, and nearly equal in size.
Only a detached right nasal is discernable in medial view (preserved upside-down; Fig. 2).It is elongated and slightly more expanded anteriorly than posteriorly. An anterior portion of the supraorbital sensory canal is enclosed in the nasal, parallel to its lateral margin.
The left antorbital is partly overlapped by the rostro-premaxillae (Fig. 2). The right antorbital, which is completely covered by the right nasal, can only be detected by X-ray scanning (Fig. 3A). Each antorbital is short and tube-like, enclosing the ethmoid canal that extends into the anterior plate of the rostro-premaxillae.
All these bones in the snout region are smooth on the surface.
Skull roofThe skull roofing bones include the fused frontals, a pair of parietodermopterotics, and three extrascapulars (Figs. 2-4). A large median ossification of the fused frontals forms most of the skull roof, which is 3.3 times as long as the parieto-dermopterotic(Fig. 2). The anterior half of this ossification is narrow, smooth, and greatly constricted above the orbital center, and its posterior half is broad and rhombic, strongly ornamented with ridges and tubercles. As revealed by X-ray scanning (Fig. 3B), a pair of supraorbital canals is enclosed in the fused frontals; each runs longitudinally through the fused frontals parallel to the lateral margin, enters the parieto-dermopterotic posteriorly and extends for a short length in this bone.
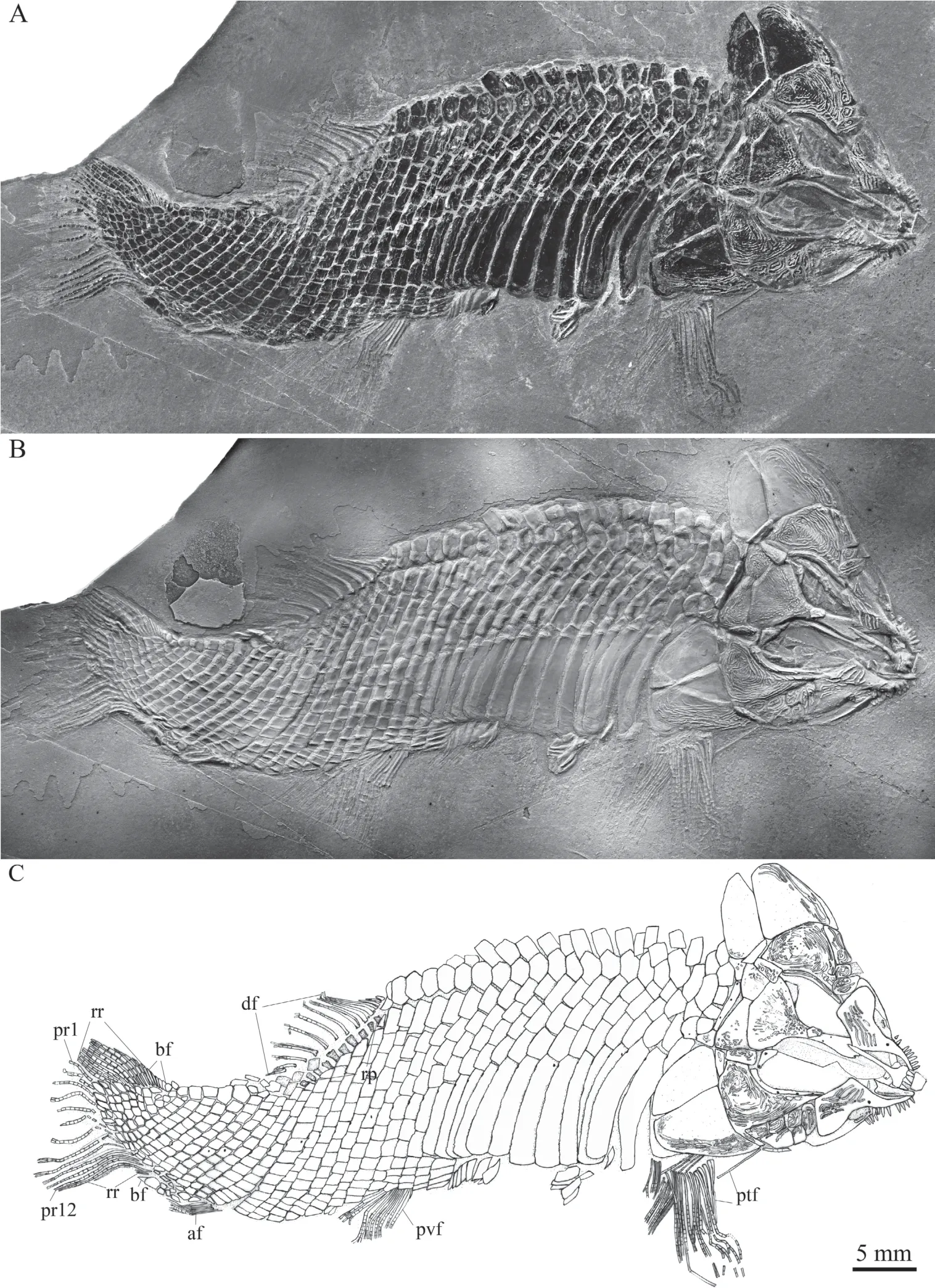
Fig. 1 Holotype of Luganoia fortuna sp. nov., IVPP V 25693
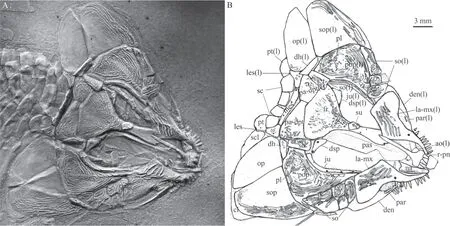
Fig. 2 Skull and pectoral girdle of Luganoia fortuna sp. nov., IVPP V 25693
The parietal is completely fused with the dermopterotic. Each parieto-dermopterotic is nearly trapezoidal with a short anterolateral extension contacting the dermosphenotic anteriorly and the posterior part of the fused frontals medially (Figs. 2, 3). In size, the paired parieto-dermopterotics are asymmetrical in the holotype; the right parieto-dermopterotic is 1.4 times as wide as the left one. Both parieto-dermopterotics taper medially and contact each other in a straight suture. Three pit-lines originate at the posterolateral area of each parietodermopterotic: the anterior and middle pit-lines are short, extending anteriorly and laterally,respectively; the posterior one is slightly longer, extending medially to reach the middle portion of this bone (Fig. 3C).
The extrascapular series includes a pair of lateral extrascapulars and a median one (Figs. 2,3). The lateral extrascapulars are small and nearly triangular. The median extrascapular is broad and anteroposteriorly short, having a convex anterior margin and a straight posterior margin.The supratemporal canal extends from the posterolateral corner of the parieto-dermopterotic into the lateral extrascapular, runs longitudinally through this bone and enters the posttemporal posteriorly. Additionally, the supratemporal commissural canal runs transversely through the middle portions of all three extrascapulars, as well revealed by the X-ray scanning (Fig. 3A).
Circumorbital bonesThe supraorbital series is incompletely preserved in the holotype with only a single detached element discernable (Fig. 2). It is small and trapezoidal,overlapping the middle region of the fused frontals.
The lacrimal is fused with the maxilla, indicated by its position and the infraorbital canal in this ossification. The lacrimo-maxilla has a large triangular dorsal process, whose posterodorsal margin defines the anteroventral margin of the orbit. The infraorbital canal exits from the antorbital, runs longitudinally through the lacrimo-maxilla and enters the jugal posteriorly (Fig. 3D).
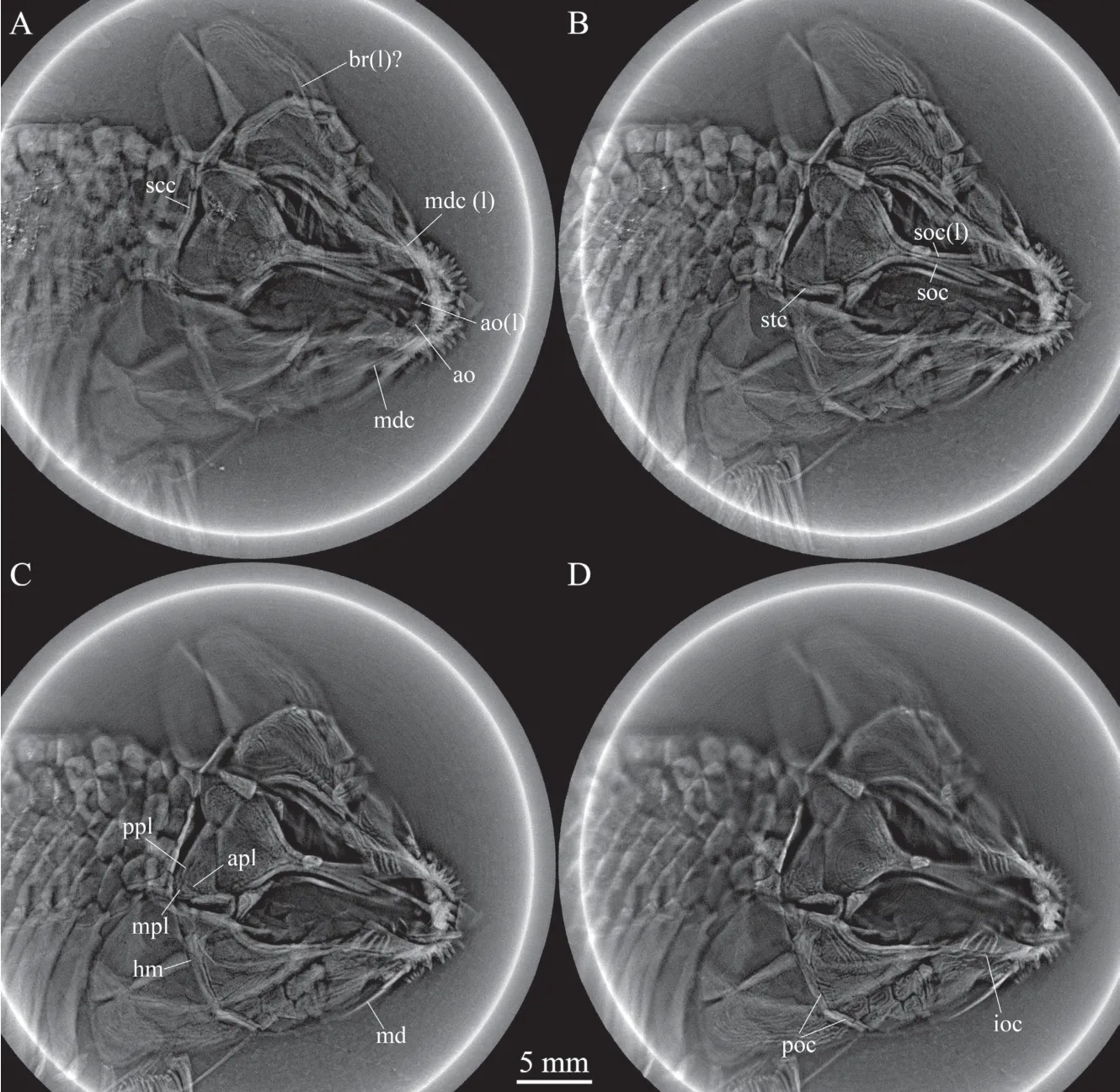
Fig. 3 Selected micro-computed scanning slices of cranial bones of Luganoia fortuna sp. nov., IVPP V 25693
The jugal is the only independent infraorbital, which is a curved, tube-like bone contacting a short posterior portion of the lacrimo-maxilla anteroventrally, the preopercle posteroventrally, and the dermosphenotic dorsally.
The dermosphenotic is small and nearly trapezoidal; it is expanded more dorsally than ventrally. This bone contacts the frontal medially and the dermopterotic and suborbital posteriorly, enclosing an L-shaped sensory canal (Figs. 2, 3C).
No sclerotic bones are discernible, and more specimens are needed to determine if these bones are present as in its European relative (L. lepidosteoides; Bürgin, 1992).
Suborbitals and dermohyalSeven suborbitals are present on the right side of the skull.They are separated into two sets by the preopercle: the dorsal set includes a single trapezoidal suborbital that inserts between the dermosphenotic and the dermohyal and lies above the preopercle. The ventral set includes six rectangular or trapezoidal suborbitals anteroventral to the preopercle. Among the ventral set, the anterior one is the deepest, while the posterior five are relatively short, arranged in two horizontal rows (three in the upper row and two below).Notably, the upper row includes only two suborbitals on the left side of the skull, displaying asymmetry in this feature (Fig. 2). All suborbitals are ornamented with ridges.
The dermohyal is a small, deep and trapezoidal bone (Fig. 2), which contacts the suborbital anteriorly, the opercle posteriorly, the preopercle anteroventrally, and the hyomandibula medially (Fig. 3C).
JawsThe premaxillae, completely fused with the rostral, have been described in the snout section. The posterior element of the upper jaw represents the fused maxilla and lacrimal. The lacrimo-maxilla is highly ornamented with ridges on its dorsally expanded posterior portion. Anteriorly, the lacrimo-maxilla has a smooth elongated ramus that contacts the antorbital dorsally and bears nine peg-like teeth on its oral margin.
Because of dorsoventral compression, the lower jaw is incompletely exposed with only parts of the dentary and prearticular discernable in medial view (Figs. 2, 3). The oral margin of the lower jaw is slightly convex and lacks a deep coronoid process. The dentary is elongated,tapering anteriorly and enclosing the mandibular canal (Fig. 3A). About ten teeth are present along the oral margin of the anterior half of the dentary. The teeth are peg-like, similar to those on the maxilla in size. The plate-like prearticular medially contacts the dentary and bears dense small conical teeth on its oral margin.
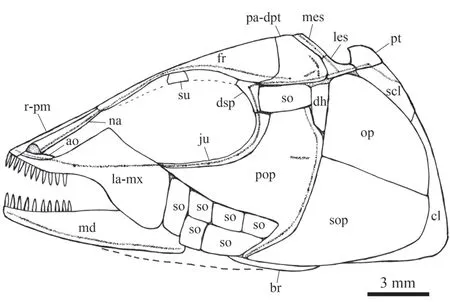
Fig. 4 Reconstruction of skull and pectoral girdle of Luganoia fortuna sp. nov., IVPP V 25693 For abbreviations see Figs. 2, 3
Operculo-gular seriesThe preopercle is large and irregular, having a broad dorsal plate, a long anterior process and a narrow ventral ramus. The anterior process inserts between the jugal and the ventral set of suborbitals, tapers anteriorly and contacts the lacrimomaxilla (Figs. 2-4). The ventral ramus of the preopercle tapers anteroventrally, contacting two suborbitals anteriorly and the subopercle posteriorly. The preopercular sensory canal extends dorsoventrally, parallel to the posterior margin of this bone (Figs. 2, 3). In addition, a horizontal pit-line is present in the dorsal plate of this bone.
The opercle is trapezoidal and nearly smooth with a depth/length radio of 0.75 (Fig. 2).The anterior and ventral margins are straight and dorsal and posterior margins convex. The subopercle is hook-shaped, one-third larger than the opercle, having a prominent anteroventral extension. Elongated and curved ridges are present only on the ventral portion of the subopercle.
The interopercle is absent as in other stem-neopterygians. No median gular is exposed,and more complete specimens are needed to determinate if it is present as in the type species of this genus (L. lepidosteoides).
Palatine and hyoid archThe parasphenoid is partly discernable in dorsal view; it is an elongated element extending longitudinally through the orbit (Fig. 2). A right hyomandibula and a possible left branchiostegal ray can only be detected by X-ray scanning. The hyomandibula is posteroventrally inclined, having a large triangular dorsal portion, a deep ventral shaft and a small posterior process that articulates with the opercle (Fig. 3C). A possible left branchiostegal ray is overlapped by the subopercle and preopercle (Fig. 3A). It is slender and slightly curved,tapering posteriorly. No other elements are discernable in the palatine or hyoid arch.
Paired girdles and finsA posttemporal, a supracleithrum, and a cleithrum are present on each side of the pectoral girdle (Fig. 2). The posttemporal is sub-circular and relatively narrow. The left posttemporal is separated from contact with the right one by three scales. The supracleithrum is anteriorly inclined, half as deep as the opercle. The cleithrum is large and curved, with its anterior portion overlapped by the opercle and subopercle. A sensory canal is enclosed in the lateral portion of the posttemporal. This canal extends posteroventrally into the supracleithrum, runs through the mid-dorsal portion of this bone and enters the first lateral line scale posteriorly. A postcleithrum is absent as in the type species of this genus.
The pectoral fins insert low on the body, and each is composed of about 15 distally segmented rays. The first is unbranched and preceded by two basal fulcra. The remaining rays are branched distally.
The pelvic girdles are not exposed. The pelvic fins insert at the 14thvertical scale row, and each is composed of nine distally segmented and branched rays that are preceded by a short basal fulcrum.
Median finsThe dorsal fin originates above the 20thvertical scale row. It is relatively long-based, composed of 11 distally segmented rays, each articulating with a radial support(Fig. 1). The first ray is unbranched and preceded by three basal fulcra; the remaining rays are branched distally.
The anal fin originates below the 23rdvertical scale row. It is very small and is composed of six distally segmented rays (Fig. 1). The bifurcations of the rays are not clear due to incomplete preservation.
The abbreviated heterocercal caudal fin is incompletely preserved at its distal portion(Figs. 1C, 5A), and it likely has a truncate posterior profile as in the type species of this genus (L. lepidosteoides; Fig. 5B). There are 12 principal rays; the marginal principal rays are unbranched, and the rest branched distally. In addition, there are ten rudimentary rays and five basal fulcra preceding the dorsal marginal principal ray. The dorsal marginal principal ray and all these rudimentary rays are located at the epaxial portion of the caudal fin. Moreover,there are three hypaxial rudimentary rays and a basal fulcrum preceding the ventral marginal principal ray. All rays are smooth on the surface.
Fringing fulcra are evidently absent in all fins.
ScalesMost of the body is covered with rhombic scales, except a small narrow region at the base of the dorsal fin, in which the body is naked (Fig. 1). The scales are smooth, arranged in 33 vertical rows between the pectoral girdle and the caudal inversion. In the 19thvertical row, there are five and eight scales above and below the lateral line scale on each side of the body. In addition, there are about ten inverted vertical rows of scales posterior to the hinge line in the caudal region, and seven scales are present in the last inverted vertical row. There are 21 dorsal ridge scales between the median extrascapular and the dorsal fin; except two relatively small scales at both tips, they are broad and hexagonal, with a straight or slightly concave posterior margin. The 13 lateral line scales in the anterior flank region are greatly deepened,each being 2.7-3.1 times as deep as the scale just above it. The 13thlateral line scale contacts two relatively short scales in the 14thvertical row, which further correspond to three horizontal rows of short scales in the mid-posterior flank region (Figs. 1, 5A). The posterior margins of the later line scales and some adjacent scales between the pelvic and anal fins are slightly serrated, and those of other scales are smooth. Each lateral line scale has a small notch at its posterior margin. Moreover, a small pore is present in some lateral line scales.

Fig. 5 Reconstruction of Luganoia
4 Discussion
The new species is referred toLuganoiaBrough, 1939 because it possesses five uniquely derived features of the genus: 1) fusion of the median rostral with premaxillae, bearing long and slender posterior process; 2) frontals completely fused into single ossification and greatly constricted above orbital center; 3) presence of a lacrimo-maxilla with a large triangular dorsal process and a concave oral margin, ending below the orbital center; 4) separation of suborbitals into two sets by a broad preopercle; and 5) presence of a horizontal row of 13 greatly deepened lateral line scales in the anterior flank region corresponding to two or three horizontal rows of short scales in the middle flank region.
In addition,Luganoiapossesses several other derived features rarely seen in other stemneopterygians:
(1) Presence of paired parieto-dermopterotics. Most stem-neopterygians have a pair of distinct parietals and dermopterotics.InLuganoia, however, the parietal is fused with the dermopterotic, resembling several other stem-neopterygians, e.g., the perleididEndennia(Lombardo and Brambillasca, 2005), thoracopterids (Griffith, 1977; Tintori and Sassi, 1992;Xu et al., 2012, 2015b) and a largeColobodus-like taxon from the Middle Triassic Luoping Biota (personal observation). This fusion of the parietal with the dermopterotic is well indicated by the presence of three pit-lines and a supratemporal canal in a single ossification.
(2) Presence of a narrow naked region of the body at the base of the dorsal fin. In adult individuals, most stem-neopterygians (except the naked thoracopterids) have their body fully covered with rhombic scales.Luganoiais unique among stem-neopterygians in having a narrow naked region on the bodyat the base of the dorsal fin. A similar condition is otherwise present in some macrosemiid ginglymodians with a long-based dorsal fin (Bartram, 1977;Schr?der et al., 2012).
(3) Absence of postcleithra. As in the type speciesLuganoia lepidosteoides,L. fortunahas both the supracleithrum and cleithrum posteriorlycontacting the first, greatly deepened lateral line scale, and lacks any postcleithra. This condition has not been known in any other early neopterygians. Two (or three occasionally) postcleithra are commonly present in Middle Triassic holosteans (Xu and Wu, 2012; Xu and Ma, 2018; Xu, 2019) and stem-teleosts (Arratia,2013; Tintori et al., 2015), but they are lost in some derived teleosts (e.g., gonorynchiforms and siluriforms).
(4) Presence of a small anal fin closer to the caudal fin than to the pelvic fin.Luganoiahas a small anal fin no larger than the pelvic fin, composed of only six or seven rays, and it is located closer to the caudal fin than to the pelvic fin. Such an anal fin is rarely seen in other Middle Triassic stem-neopterygians with greatly deepened scales in which the anal fin is commonly more anteriorly placed and larger than the pelvic fin (Bürgin, 1992; Lombardo,1999; Mutter and Herzog, 2004; Sun et al., 2012, 2015).
Notably, the discovery ofLuganoia fortunareveals several phylogenetically important features of the genus: the presence of a pair of antorbitals (unknown inL. lepidosteoidesdue to incomplete preservation); absence of a deep coronoid process of the lower jaw (contra Brough,1939 but consistent with Bürgin, 1992); and absence of a plate-like quadratojugal (the previously alleged ‘quadratojugal’ inL. lepidosteoideslikely represents a ventral preopercular element).
Moreover, the discovery ofL. fortunaprovides new information on the morphological diversification of the genus. The new species is easily distinguished from the type species of the genus (L. lepidosteoides) by the following features: 1) relatively small parietodermopterotic (frontals 3.3 times as long as parieto-dermopterotics vs. 2.2 times inL.lepidosteoides); 2) three extrascapulars (four inL. lepidosteoides); 3) single suborbital above preopercle (two inL. lepidosteoides); 4) six or seven suborbitals below preopercle (five inL. lepidosteoides); 5) subopercle 1.5 times as large as opercle (subopercle nearly as large as opercle inL. lepidosteoides); 6) preopercle with an anteroventrally tapering ramus (absent inL. lepidosteoides); 7) 11 dorsal fin rays (14 inL. lepidosteoides); 8) anal fin smaller than pelvic fin (nearly equal inL. lepidosteoides); 9) 12 principal caudal fin rays (14 inL. lepidosteoides);10) three teeth on the rostro-premaxillae (four or five inL. lepidosteoides); and 11) three scales between posttemporals (single inL. lepidosteoides).
Furthermore, the discovery ofL. fortunais of special interest in paleobiogeography,extending the geological distribution of the genus from the latest Anisan to Ladinian of the southern and eastern Switzerland, northern Italy and Catalonia in Spain (a part of the western Paleotethys Ocean) into the Ladinian of Xingyi, Guizhou in South China (a part of the eastern Paleotethys Ocean). The first occurrence of the type speciesL. lepidosteoidesin the latest Anisian indicates that the genus originated no later than ~242 Ma, about two million years earlier thanL. fortuna. The original center ofLuganoiais less easy to determine. Assuming that the genus originated in Europe based on the present fossil record, the Paleotethys Ocean would have provided a west-east corridor for the dispersal of this genus into South China.L.fortunadocuments the first record of the Luganoiidae in China (or even Asia). Although the short and deep bodiedBrachysomusfrom the same fossil assemblage was previously referred to the Luganoiidae (Liu et al., 2003), this is based on inaccurate anatomical identifications.My comparative studies of the holotype and new specimens collected in recent years indicate thatBrachysomusis a ginglymodian within the neopterygian crown. A detailed revision ofBrachysomusis out of the scope of this paper and will be presented in the future.
5 Conclusion
The discovery ofL. fortunaprovides an important addition to our understanding of the taxonomical diversity of the Middle Triassic (Ladinian) Xingyi Biota. The new finding represents the first record of the Luganoiidae in Asia, indicating that the biogeographical distribution of this family is much wider than previously recognized. Comparative studies of the Chinese species with the slightly older type species from Europe reveal some anatomical features previously unnoticed inLuganoia, and an amended diagnosis of the genus is presented. Additionally, the study specially emphasizes the distinguishable features between the Chinese and European species, and the results provide valuable information on the morphological diversification ofLuganoia.
AcknowledgmentsI thank Chang M.-M. and Bürgin T. for valuable comments on a previous version of this manuscript, Yin P.-F. and Lin H.-X. for conducting X-ray computed tomography, Furrer H. for access to comparative fossil material in the Pal?ontologisches Institut und Museum, Uinversit?t Zürich, and Li N. for helps in fossil collection.
 古脊椎動(dòng)物學(xué)報(bào)(中英文)2020年4期
古脊椎動(dòng)物學(xué)報(bào)(中英文)2020年4期
- 古脊椎動(dòng)物學(xué)報(bào)(中英文)的其它文章
- Ancient DNA molecular identification and phylogenetic analysis of Cervinae subfossils from Northeast China
- New skulls of ctenodactyloids from the Early Oligocene of Ulantatal, Nei Mongol, China
- Taxonomic revision of Anthracokeryx thailandicus Ducrocq, 1999 (Anthracotheriidae, Microbunodontinae)from the Upper Eocene of Thailand
- New chroniosuchian materials from Xinjiang, China
- Subdivision and age of the Silurian fish-bearing Kuanti Formation in Qujing, Yunnan Province
- 總 目 次